Zhan Wang1,
Peng-yuan Zheng2 ![]()
For correspondence:- Peng-yuan Zheng Email: zhengpengyuan765@hotmail.com Tel:+8637166916801
Received: 30 November 2015 Accepted: 15 October 2016 Published: 28 November 2016
Citation: Wang Z, Zheng P. Apigenin inhibits proliferation and migratory properties of Barrett esophageal adenocarcinoma cells by targeting PI3K/Akt/mTOR pathway. Trop J Pharm Res 2016; 15(11):2365-2370 doi: 10.4314/tjpr.v15i11.10
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the effect of apigenin on Barrett's esophagus–associated esophageal adenocarcinoma (BEAC) cells OE33, and also to ascertain the mechanism by which it inhibits cellular proliferation and motility.
Methods: Proliferation index of OE33 in the absence and presence of apigenin was determined by methyl-thiazolyl-tetrazolium (MTT) assay and apoptosis was determined by enzyme-linked immunosorbent assay (ELISA) method. Boyden Chamber’s assay was applied to determine the migration and invasion of control and apigenin-treated OE33 cells. Status of PI3K/Akt/mTor signaling was further determined by Western blotting in control and apigenin-treated cells.
Results: Apigenin resulted in the inhibition of the proliferation of OE33 cells in a dose- and time-dependent fashion, with an IC50 of 75 µM, after 72 h of incubation, and also induced apoptosis, with modulation of pro- and anti-apoptotic genes. Furthermore, apigenin inhibited the motility of OE33 by targeting PI3K/Akt/mTOR signaling.
Conclusion: Apigenin effectively inhibits the oncogenicity of OE33 cells by targeting PI3K/Akt/mTOR pathway.
Introduction
Apigenin [5,7-dihydroxy-2-(4-hydroxyphenyl)-4H-1-benzo-pyran-4-one] is a dietary flavone, found abundantly in fruits and variety of vegetables including parsley, oranges , onions, tea, chamomile, wheat sprouts, etc [1,2]. Apigenin has immense therapeutic potential due to its anti-inflammatory, antiviral, antimutagenic as well as anticancer properties [2-10]. In certain cancer cells, apigenin has been known to target the PI3K/Akt/mTOR signaling leading to inhibition of oncogenicity [11-14].
Barrett's esophagus–associated esophageal adenocarcinoma (BEAC) has become the fastest growing tumor in the western world, within the last three decades [15,16]. In the United States alone, the incidence of BEAC had increased by 300 % to 500 % in the last 40 years [17] and recently published meta-analysis studies reveal that similar risk factors of Barrett's esophagus also prevail in Asian countries [18]. Barrett’s esophagus (BE) is known to be the most significant precursor lesion for esophageal adenocarcinoma (EAC) [19], which resulted from the longstanding exposure of the esophageal mucosa to gastro-esophageal reflux, resulting in the metaplastic replacement of the native squamous epithelia by columnar cells (i.e. Barrett’s mucosa) [20,21]. This epithelial population serves as the platform for the formation of intra-epithelial neoplasia, also termed as dysplasia, which thereafter might lead to the Barrett-related EAC [20,21].
Since the effect of apigenin on BAEC is not investigated previously, we conducted this research to find out whether apigenin can inhibit BAEC. For our study we have used esophageal adenocarcinoma cell line OE33, as the in vitro model and also investigated the mechanism of apigenin action on OE33 cells.
Methods
Materials
Apigenin was purchased from Sigma. Nutrient mixture Dulbecco’s minimal essential medium (supplemented with 1 mM L-glutamine), fetal bovine serum, penicillin-streptomycin, and 0.25 % Trypsin-EDTA were purchased from GIBCO (Invitrogen). Primary antibodies such as Bax, and Bcl-2, were purchased from Santa Cruz Biotechnologies (Santa Cruz, CA, USA), whereas cleaved caspase-3, cleaved caspase-9,PI3K (p85), p-Akt ( Ser473), and p-mTor (Ser2448) were purchased from Signaling Technologies (Beverly, MA, USA).
Cell culture and maintenance
BEAC cell line OE33, was purchased from American Type Culture Collection (ATCC). OE33 cells used in this study were from fifth through tenth passages and were routinely maintained in Dulbecco's Modified Eagle's Medium supplemented with 10 % FBS and antibiotics , penicillin (100 U/mL), and streptomycin (100 μg/mL), at 37 °C in a humidified chamber.
Cell viability assay (MTT assay)
Cultured OE33 cells were seeded in 96 well plate at a density of 1 × 104 cells/ml and grown to confluency. Cell were then treated with different concentrations of apigenin (0-100 µM) for 24, 48 and 72 h, respectively. After treatment, MTT solution was added to each well and the absorbance was measured on an ELISA reader (MultiskanEX, Lab systems, Helsinki, Finland) at a wavelength of 570 nm according to published protocol (29). Data are presented as percent inhibition, calculated as in Eq 1.
Inhibition (%) = {100-(At-As)/100} …………… (1)
where At and As are the absorbance of the test substances and solvent control, respectively.
Apoptosis assay
Involvement of apoptosis in apigenin treated OE33 cells was determined using the apoptosis detection ELISA kit (Roche Diagnostic, USA) following. Briefly, control and apigenin-treated cells were subjected to lysis and centrifuged at 1500 x g for 15 min. The cytosolic supernatant was collected, and the total protein was estimated. Around 10 μg of protein, from each sample was added to streptavidin-coated well, containing 80 μL of reaction mixture consisting of anti-histone-biotin and anti-DNA–peroxidase and incubated for 2 h with continuous shaking. Finally the chromagenic substrate ABTS (2,2¢-azino-di-(3-ethylbenzthiazoline-6-sulfonic acid)) was added into each well to get a colored product, which was measured by the ELISA reader at 405 nm.
Determination of migration/invasion by Boyden Chamber assay
Migratory properties of OE33 cells were determined by Boyden chamber assay, using track-etched polyethylene terephthalate membranes (Corning, MA), with 8.0 μm diameter-pore size. OE33 cells were treated with apigenin (0 – 75 µM) for 48 h, and migration assay was performed accordingly. Invasion was performed following the published protocol, using Matrigel®-coated invasion chambers (BD Biosciences). The cells which migrated/ invaded to the lower surface of the membrane were stained with crystal violet. The resultant crystal violet complex was then dissolved in 10 % acetic acid and the absorbance was measured at 600 nm, using the microplate reader (Multiskan EX, Lab systems, Helsinki, Finland) to determine the extent of migration or invasion.
Western blot analysis
Cultured OE33 cells after treatment with apigenin, were extracted in ice cold lysis buffer, constituted of 150 mM NaCl, 20 mM Tris-HCl, 1 % NP-40, 20 μg/mL leupeptin, 20 μg/mL aprotinin, 1 mM ortho-vanadate, and 2 mM PMSF, pH 7.4. Total protein from the cell lysates were estimated by Bradford method. Approximately, 30 - 50 μg of protein from each sample was separated by 10 - 12 % SDS-PAGE, electrophoretically transferred to polyvinylidene difluoride membrane, incubated with different monoclonal and polyclonal antibodies according to manufacturer’s mentioned dilution and subsequently with required secondary antibodies. Then, the membranes were exposed to Kodak x-ray films after chemiluminescent treatment.
Statistical analysis
Analysis of data was achieved with GraphPad Prism version 4.00 (GraphPad Software, San Diego, California, USA). All data are expressed as mean ± SD. Statistically significant differences between groups were determined by paired Student's two-tailed t-test. P < 0.05 was considered statistically significant.
Results
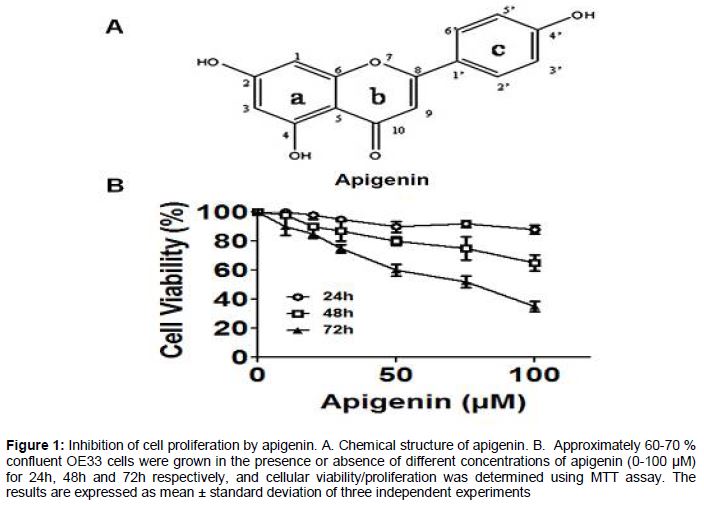
Apigenin inhibits proliferation of OE33 cells
Treatment of esophageal adenocarcinoma cells, OE33 with apigenin resulted in the decrease of cellular proliferation in both concentration and time-dependent fashion (B). OE33 cells were treated with different concentrations of apigenin (0 - 100 µM) for 24, 48 and 72 h, respectively. A drastic reduction in the cell viability was observed after 72 h of treatment and the IC50 was observed at a concentration of 75 µM apigenin ().
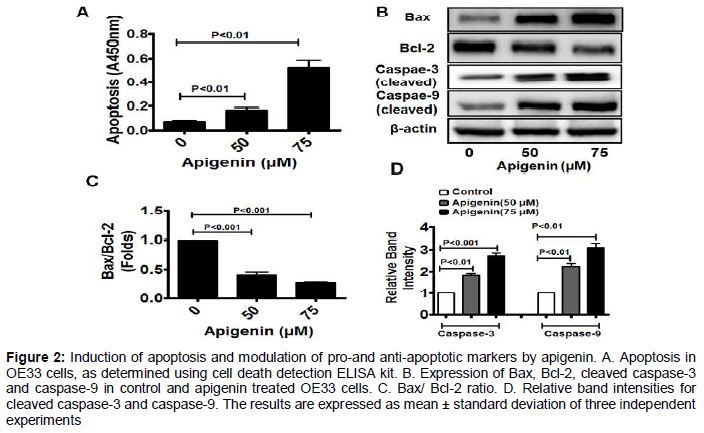
Apoptosis and modulation of pro-and anti-apoptotic markers
To find out the mode of cell death in apigenin-treated OE33 cells we determined the involvement of apoptosis. In the presence of 50 µM apigenin apoptosis was increased by 2.4-fold, while in presence of 75 µM apigenin, it was increased by 7.5-fold. Thus the results indicate that apigenin can induce apoptosis in OE33 cells in a concentration -dependent fashion (A). Moreover we also observed the status of Bax/Bcl-2 ratio in apigenin-treated OE33 cells. We observed that, the pro-apoptotic protein Bax is up regulated, while the anti-apoptotic protein Bcl-2 is down regulated, the increasing concentrations of apigenin (B and C). Also significant up regulation of cleaved caspase-3 and caspase-9 were observed, suggesting that apigenin is activating the intrinsic apoptotic pathway in OE33 cells (D and 2E).
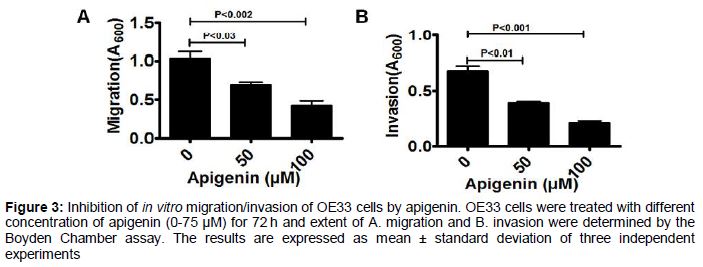
Apigenin inhibits the motility of OE33 cells
Migratory or invasive properties of aggressive cancer cells are the hallmark of tumor metastasis. To find out whether, apigenin can inhibit the motility of OE33 cells we performed the migration as well invasion assays using the Boyden Chamber (). Significant inhibition of cellular migration was observed at 50 µM apigenin (~33 %), while in presence of 75 µM apigenin, the migration was reduced by almost 60 % (A). Similar pattern was observed when invasion assay was performed with apigenin-treated cells and the invasive phenotype of OE33 cells were inhibited by apigenin in a concentration dependent fashion (B).
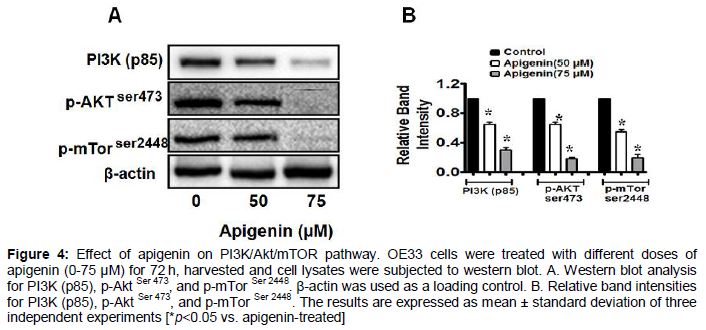
Apigenin inhibits the PI3K/Akt/mTOR pathway in OE33 cells.
PI3K/Akt/mTOR signaling plays vital role in cell proliferation, growth and survival and is one of the pathways frequently activated in cancer, including BEAC [17-19]. Many anticancer agents act by down regulating this signaling pathway, thus promoting apoptosis in cancer cells (15, 16). To further explore the mechanism of cell death caused by apigenin, the expression and phosphorylation status of the components of this signaling pathway were analyzed (). We observed that, apigenin- treatment caused significant reduction in p85, the regulatory subunit of PI3K and also reduced the expression of phosphorylated (Ser 473) form of Akt. The effect of apigenin on mTOR activity was further investigated and exposure of OE33 cells to apigenin resulted in decrease in the level of activated form of mTOR (phosphorylated form- Ser2448), although the total Akt and mTOR levels remained unaffected by the treatment (data not shown).
Discussion
The beneficial health effects of dietary phytochemicals have made them promising candidates for treatment and prevention of multiple diseases, including cancer. However, precise cellular and molecular targets for dietary supplements remain largely unknown. Apigenin is a dietary flavone, commonly associated with fruit- and vegetable-diets [1,2]. This dietary flavonoid has potential anticancer properties, but most importantly it has negligible adverse effects on noncancerous cell types. There was no earlier report of the mode of action of apigenin on esophageal adenocarcinoma or EAC. Hence in the present study we have investigated in detail, the mechanism by which apigenin imparts its growth inhibitory effect on EAC cells, OE33.
It was observed that apigenin exhibits superior anticancer properties by inhibiting the cellular proliferation of OE33 cells in both concentration and time-dependent fashion. Further experimental evidences confirmed that, this dietary flavonoid is effective in inducing apoptosis in OE33, in a concentration dependent fashion. Expectedly the Bax/Bcl-2 ratio was found to increase with increased apigenin concentration, confirming the involvement of apoptosis in apigenin mediated cytotoxicity. Furthermore, it was observed that apigenin-induced apoptosis is mediated by caspase-3 dependent pathway. Since BEAC is highly metastatic in nature, we determined whether apigenin can inhibit the motility of OE33 cells. Very interestingly, we observed that, apigenin-treatment resulted in the inhibition of both migratory as well as invasive properties of OE33 cells in a concentration dependent fashion.
Considering the important role of PI3K/Akt/mTor signaling in the advancement of BEAC, we investigated whether apigenin can block this pathway in OE33 cells. Our western blot data confirmed that, apigenin targets this oncogenic signaling pathway in OE33cells. Expression levels of p85, the regulatory subunit of PI3K, were drastically down regulated by apigenin in a concentration dependent fashion. Moreover the phosphorylation-dependent activation of the downstream modulators, such as p-Akt (Ser-473) and p-mTor (Ser2448) were also inhibited by apigenin treatment.
Conclusion
The findings of the present study serve to delineate the anticancer mechanism of the dietary flavone, apigenin, against BEAC cells, which has the potential of a major therapeutic regimen against esophageal adenocarcinoma.
Declarations
Acknowledgement
References
Archives


News Updates